Invited Talk 3
Thursday, July 21, 9:00-9:50 (International Conference Hall)
Brain-Body-Environment Interactions in Cognition
Toshio Inui
Kyoto University, Japan
Abstract
1. The visual
world and estimation of the outer world
The visual world is estimated from retinal images and represented in
the brain. We thus conduct ourselves in our own world. In order to behave successfully
in such a visual world in the brain, the world must be stable and sensitive
to various changes. In addition, when dealing with moving objects, we must be
able to predict states beforehand. We can therefore control our own actions;
and our visual world must also be appropriate for this purpose. In actual fact,
we take advantage of such control using predictions when we control our own
body movements.
2. Two visual systems and embodied cognition
The study published by Goodale in 1991 has exerted a great deal of
influence on the brain and cognitive sciences. He discovered that two functions
are generated by different pathways in the brain; one is the function for object
manipulation; the other, for object recognition. Neural activities from the
occipital lobe to the parietal lobe (the dorsal system) are responsible for
the utilization of knowledge about how to use objects, known as procedural or
functional knowledge. Conversely, activities from the occipital lobe to the
temporal lobe (the ventral system) are essential to utilizing knowledge about
the classification of shapes or naming of objects.
The dorsal system is involved in processing information about how
to use objects, and is a “perception for action system”. When we
try to drink from a glass of water in front of us, we reach out our arm and
grasp the glass. To achieve this, we must accurately gauge the spatial relationship
between our hand and the object (e.g., the position of the glass compared to
the center of our body). We must also realize the appropriate plan to move our
hand and arm. Nevertheless, such action is typically completed without any difficulties
whatsoever. We seem to follow the requisite procedure automatically and unconsciously
at the same time as looking at the glass. Therefore it can be said that we perceive
an object not only with vision, but also with our body. Such a cognitive function
is called “embodied cognition”, and results from good coordination
between vision and motor abilities, based on close communication among the occipital
and parietal lobes and the motor system.
The function of such a mechanism of coordination brings about not
only the transformation of visual information into motor information, but also
the generation of visual images from a motor data set. Thanks to smooth execution
of these bidirectional processing, we perform various mental simulations in
the brain. In fact such mental simulation is extremely important for the function
of the mind. Furthermore, a similar mechanism is considered to exist between
auditory perception and articulation. As we are capable of learning through
imitation with visuo-motor coordination, we can also learn articulation with
auditory-motor coordination. This is important in learning a second language,
as well as the native language.
3. Brain-body-environment
We skillfully perform various manipulations of the environment or information
obtained from the environment. Most of these manipulations go succeed with predictive
processing of information. Assessing whether a manipulation will result in the
expected state is prerequisite for predictive processing or predictive control.
We therefore receive sensory feedback from the environment or body and compare
this with the predicted state. This indicates the importance of both visual
and somatosensory information, including tactile information. Merleau-Ponty
compared relationships between humans and the environment as described above
to exchanges between a subject and an object, or exchanges between something
that touches and something that is touched. Complex interrelations should develop
in such situations, rather than a simple and static relationship between the
subject and object. Thus, the subject who perceives the environment is considered
to be involved in an inseparable and complex relationship with the environment.
4. Prediction and imitation
The imitation and prediction of actions by other organisms represent the basis
for creating high-level cognitive functions. To imitate the actions or voices
of other people, visual or auditory information must be transformed into self-generated
movements. In the same way as an infant imitates the parents, parents imitate
infants. These interactions are connected through dialogue with gestures. Melzoff
and Gopnik (1993) proposed that the infant extends the cognitive functions through
such “mutual imitation games”. The important problem here is that
an infant understands the equivalence of information from different modalities
(visual information on action by others and motor information by the self).
Another issue involves the mechanisms used to correct error of imitation. Melzoff
and Moore (1977) suggested a conceptual model for these problems. However, much
more research is needed before a clear understand of this topic is achieved.
The mirror neuron, discovered by Rizzolatti et al. (1996), is clearly
involved in comprehension of the actions of others. Moreover, Arbib and Rizzolatti
(1996) have suggested that the transformation from visual information to movement
is realized in the pathway from STS to F5, whereas the transformation from movement
to visual information is conducted in the pathway from F5 to STS. They proposed
that these pathways play important roles in comprehending the gestures of others.
5. Investigating neural correlations of state estimation
in visually guided movements
State estimation of self-movement, which is based on both motor commands and
sensory feedback, has been suggested as the essence of human movement control
to compensate for inherent feedback delays in sensorimotor loops. We have investigated
the neural basis for state estimation of human movement (Ogawa et al., 2005)
using event-related functional magnetic resonance imaging (fMRI).
Participants performed visually guided movements with an artificial
delay introduced into actual visual feedback during movement. As movement control
based purely on feedback information is impaired under delayed visual feedback
conditions, prediction of self-movement was considered likely to be used to
maintain relatively accurate performance in the presence of feedback delays.
Such internal state estimation was then compared with actual feedback information,
and prediction error was used to optimize the estimated state. This study investigated
the neural basis of state estimation and prediction errors of self-movement
by analyzing correlations between motor performance and brain activity under
conditions of delayed visual feedback. Behavioral data were also measured during
the task and used as explanatory variables in analysis of event-related fMRI.
Such parametric fMRI analysis based on motor performance of each subject could
reveal relationships between motor performance and brain activity.
Experiment 1 investigated brain activations that were significantly
correlated with visual feedback delay and motor error by parametrically manipulating
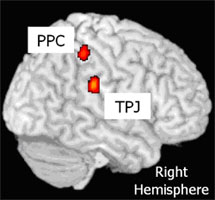
visual feedback delay. Activation of the right posterior parietal cortex (PPC)
was positively correlated with motor error, whereas activation of the right
tempo-parietal junction (TPJ) was observed only in the group with a smaller
increase in motor error following increased visual feedback delay. Experiment
2 involved parametric analysis of motor performance while controlling computer
mouse movement speed during the task. Activity in the right TPJ displayed a
significant positive correlation with motor performance under delayed visual
feedback conditions. In addition, activity of the PPC was greater when motor
error was presented visually (Fig. 1).
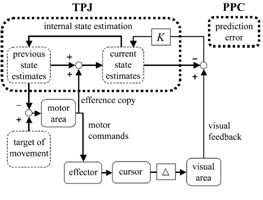
The relationship between activated regions in this study and the
predictive control model are shown schematically in Figure 2. First, a motor
control model is generated in the central nervous system (CNS) and sent to the
effector. Actual movement is then generated and visual feedback is received
as the cursor position on a display. Simultaneously, the efference copy of motor
commands is sent to the CNS, which internally estimates self-movement using
the copy and previous state estimates as inputs. State estimates are used to
generate the next motor commands in order to compensate for external feedback
delay (?). Finally, state estimates are compared with actual feedback information,
and prediction error is used to optimize state estimates, multiplied by the
appropriate observer gain (K). The current results suggest that the PPC is involved
in such a prediction error with regard to visual coordinates, and that the TPJ
is involved in state estimation of movements.
6. Tool-use, pantomiming and imagination
Neuropsychological studies have shown that some patients with ideomotor apraxia
(IMA), typically due to left parietal lesions, are unable to pantomime with
tools, despite the ability to manipulate the actual tools normally (Rapcsak
et al., 1995; De Renzu and Lucchelli, 1988). This finding suggests that the
neural mechanism for manipulating tools in everyday life differs from that for
pantomiming tool use.
We examined the neural basis underlying tool-use behavior from the
perspective that use of a familiar tool involves a specific pathway that generates
actions without explicit retrieval of intended actions (Imazu et al., 2005).
Regional cerebral blood flow in healthy Japanese subjects was measured using
fMRI while the subject picked up an object with chopsticks, picked up an object
with the hand, pantomimed the use of chopsticks, imagined the use of chopsticks,
and imagined use of the hand. When comparing activations while picking up an
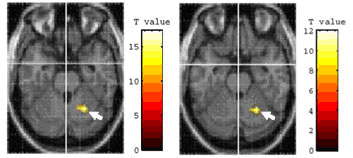
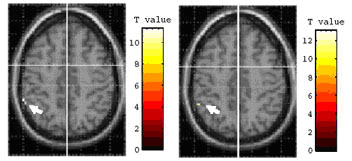
object using chopsticks with activations while imagining the use of chopsticks
(TI), bilateral areas of the cerebellum and the primary motor area were significantly
activated (Fig. 3a).
Using inverse contrast, TI versus TU, the left inferior parietal
lobule (IPL) was significantly activated (Fig. 3b). Comparison of TU and pantomiming
the use of chopsticks (Tp) revealed significant activation in the lateral right
cerebellum and bilateral postcentral gyri (Fig. 3a). Using inverse contrast,
Tp versus TU, significant activation in the left inferior parietal lobule was
observed (Fig. 3b).
Given these results, the left IPL appears to selectively contribute to tasks
requiring explicit retrieval of tool-related hand movement, and pantomiming
and imagining tool use. Furthermore, a comparison of these tasks revealed that
activation in the lateral part of the right cerebellum increased during execution
of tool-use. The present findings support our speculation that actual use of
a familiar tool involves a different pathway from that of the mental imagery
process.
Acknowledgment
This study was supported by (1) the 21st Century COE Program, (2) the Advanced
and Innovational Research Program in Life Sciences from Ministry of Education,
Culture, Sports, Science and Technology, the Japanese Government, and (3) the
Research for the Future Program administered by the Japan Society for the Promotion
of Science (JSPS-RFTF99P01401).
References
[1] Arbib, M.A., and Rizzolatti, G. (1996) Neural expectations: a possible evolutionary
path from manual skills to language. Communication and Cognition, 29, 3, 393-424.
[2] De Renzi, E., and Luchelli, F. (1988) Ideational apraxia. Brain, 111, 1173-1185.
[3] Goodale, M.A., Milner, A.D., Jakobson, L.S., and Carey, D.P. (1991) A neurological
dissociation between perceiving objects and grasping them. Nature, 349(6305),
154-156.
[4] Imazu, S., Sugio., T and Tanaka., S and Inui, T. (2005) The difference between
actual usage of chopsticks and imagining of chopsticks use: an event-related
fMRI study. Cortex. in press.
[5] Meltzoff, A., and Gopnik, A. (1993) The role of imitation in understanding
persons and developing a theory of mind. In Baron-Cohen, S., Tager-Flusberg,
H. and Cohen, D.J. (Eds.), Understanding other minds, 335-366. New York: Oxford
Medical Publications.
[6] Meltzoff, A.N., and Moore, M.K. (1997) Explaining facial imitation: A theoretical
model. Early Development and Parenting, 6, 179-192.
[7] Ogawa, K., Inui, T., and Sugio, T. (2005) Neural correlates of state estimation
in visually guided movements: an event-related fMRI study. Cortex. in press.
[8] Rapcsak, S.Z., Ochipa, C., Anderson, K.C., and Poizner, H. (1995) Progressive
ideomotor apraxia, evidence for a selective impairment in the action production
system. Brain and Cognition, 27, 213-36.
[9] Rizzolatti, G., Fadiga, L., Gallese, V., and Fogassi, L. (1996) Premotor
cortex and the recognition of motor actions. Cognitive Brain Research, 3, 131-141.
|
|
|
Figure.1 The posterior parietal cortex (PPC) and the temporo-parietal
junction (TPJ) observed in the current study. |
Figure.2 The predictive control model of self-movement and
brain activations observed in the current study. |
| |
Z = -26 |
Z = -24 |
Z = 46 |
Z = 46 |
|
|
TU > TP (p < 0.05,uncorrected) |
TU > TI (p < 0.05,uncorrected) |
TP > TU (p < 0.05,uncorrected) |
TI > TU (p < 0.05,uncorrected) |
a) The activation in the
right cerebellum |
b) The activation in the
left IPL |
Figure. 3 |
Profile
He is now a professor at the Department of Intelligence
Science and Technology, Graduate School of Informatics, Kyoto University. His
majors are cognitive science and computational neuroscience. Presently engaged
in research of visual information processing and the communication mechanism.
He is an executive committee member of the Neuropsychology Association
of Japan, the Japanese Society for Cognitive Psychology, the Japanese Neuro-ophthalmology
Society, and the Japan Human Brain Mapping Society. He serves on the editorial
board of Neural Networks and Japanese Psychological Research. He is also a chair
of Human Communication Group of IEICE (the Institute of Electronics, Information
and Communication Engineers), an academic counselor member of International
Institute for Advanced Studies, and an executive member of Brain Century Promotion
Conference (non profit organization). His publications include “Inui,
T., and McClelland, J. L. (Eds., 1996) Attention and Performance XVI : Information
integration in perception and communication. Cambridge, MA: The MIT Press.”
Back